|
|
Agustín P. Monteoliva * y Carlos Muñoz Bellido **
*Biólogo.
Consultor de Medio Ambiente, Infraestructura y Ecología, S.L.
**Ingeniero de Caminos, Canales y Puertos, Confederación
Hidrográfica del Duero
Descriptores: Embalse, Gestión de calidad de aguas, Integridad ecológica, Eutrofización, Limnología
Introducción
Es bien conocido que los ecosistemas limnéticos reciben y procesan la
tensión que se ejerce en el conjunto de la cuenca de drenaje (Margalef,
1983). Tanto el sistema acuático receptor como la cuenca en sí
misma (Frisell et al., 1986), se disponen en niveles de organización
jerárquica, cuya estructura y funcionalidad responden a determinadas
escalas espacio-temporales (Munkittrik, 1995).
Esta organización
jerárquica produce una respuesta reguladora ante las tensiones externas,
de forma que en cada nivel de organización se genera una nueva salida
que repercute en el nivel siguiente. Por lo tanto, se produce una transmisión,
previo procesamiento de la tensión de entrada, que genera unos efectos
determinados en cada nivel, en lapsos de tiempo y extensiones espaciales congruentes
con los procesos determinantes de la respuesta en ese nivel.
A medida que se asciende en la aludida escala jerárquica, se pierde sensibilidad
y predictibilidad, pero la respuesta que se obtiene es más integradora
y representa en mayor medida el estado real del ecosistema. Así, por
ejemplo, la consideración de determinadas comunidades de invertebrados
bentónicos permite informar sobre la repercusión global de hitos
contaminantes esporádicos que difícilmente serían detectables
en determinaciones discretas de parámetros físico-químicos;
aun cuando fueran detectables mediante programas de medición en continuo,
no informarían sobre los efectos reales sobre el ecosistema de esas tensiones
o cargas de estrés.
Sin embargo, la evaluación sistemática del estado de la calidad
de las aguas se suele circunscribir a determinados elementos que tienen relevancia
a efectos de los usos y explotación del recurso hídrico. Estos
elementos no se corresponden necesariamente con niveles de organización
en los que los procesos implicados resultan cruciales para entender y, por lo
tanto, corregir la respuesta observada.
No obstante, hay que destacar que la legislación ambiental europea está
comenzando a incorporar estos conceptos ecosistémicos. Así, por
ejemplo, la Propuesta de Directiva Marco sobre la Calidad Ecológica de
las Aguas 1
establece en su artículo 4, dedicado a los objetivos ambientales, que
"se tratará de prevenir el deterioro del estado ecológico
y la contaminación de las aguas superficiales (…) o en el caso de
las masas de agua muy modificadas y/o artificiales (embalses) un buen potencial
ecológico y un buen estado químico."
En el caso de los embalses, ecosistemas de características intermedias
entre un río y un lago, se produce una compartimentación física
que incrementa los tiempos de residencia y condiciona las salidas. De esta forma,
la impronta que se transmite al río aguas abajo de la presa depende no
sólo de las entradas externas al sistema, sino también de las
características de estos compartimentos. Dado que se trata de sistemas
más controlados que los naturales y cuyas variables físicas son
más conocidas, son mayores las posibilidades de entender los procesos
en niveles de organización superiores (biológicos y ecosistémicos)
y manejar las respuestas que se producen.
Problemática
En un país como España, con más de mil embalses, tiene
sumo interés mantener su integridad ecológica, minimizando así
los riesgos de colapso (fallos en los mecanismos de amortiguación u homeostáticos)
en el propio embalse y en los tramos fluviales de influencia. En este sentido,
la eutrofización cultural se sitúa entre las alteraciones más
extendidas y más problemáticas en los embalses, considerando además
que modifica también la capacidad de respuesta del sistema a otros tipos
de contaminación.
La eutrofización es un proceso de enriquecimiento de las aguas en sustancias
nutritivas primarias (nitrógeno y fósforo principalmente), que
si es excesivo conduce a modificaciones sintomáticas en los ecosistemas
acuáticos, tales como un aumento de la producción primaria (pelágica
en embalses), y una simplificación de la estructura de las comunidades
biológicas del sistema.
La secuencia de cambios que acontecen como respuesta a una carga excesiva de
nutrientes comienza con un notable incremento de la productividad primaria.
Este incremento es seguido por uno paralelo de los demás niveles tróficos
del ecosistema. Como consecuencia, la cantidad de materia orgánica que
se genera es mayor y los procesos oxidativos de la misma serán de mayor
envergadura.
Si, como sucede en nuestro ámbito geográfico, las capas superficiales
de la masa de agua están sometidas a un intenso calentamiento estival
que genere una estratificación térmica, la oxidación de
los restos orgánicos que se depositan en el fondo de la cubeta y la preponderancia
de la respiración sobre la fotosíntesis en las capas profundas
pueden acarrear finalmente un estado anóxico del hipolimnion.
Se trata, además, de un proceso con retroalimentación positiva,
puesto que, a medida que se incrementan los aportes, el sistema se descompensa,
pierde capacidad de asimilación de los mismos y se incrementa la recarga
interna desde los sedimentos hacia la columna de agua. Por otro lado, la producción
bacteriana se incrementa en profundidad, debido a la proliferación de
organismos anaerobios facultativos y estrictos, nitrobacterias y sulfobacterias,
en general. Estas últimas generan anhídrido sulfhídrico,
un veneno respiratorio para la fauna acuática.
Como consecuencia
del incremento del desarrollo algal hay una pérdida de transparencia
del agua, que adquiere olores y sabores desagradables y que podría padecer
toxicidad por el desarrollo de ciertas cianofíceas. La calidad del agua
se ve sensiblemente menguada, lo que dificulta y limita los posibles usos a
los que se puede destinar.
La problemática más comúnmente asociada a este proceso,
y que repercute en detrimento de las posibilidades y coste de uso del agua,
se glosa en el cuadro 1 (Landner, 1976; Vollenweider, R.A., 1989; Margalef,
1983; Odum, 1985).
 La eutrofización puede tener un origen natural, proceso lento, aunque
algo menos si se dan unas determinadas características geomorfológicas
en la cuenca (cuencos poco profundos y rocas no calizas), suelos ricos en nutrientes
y climas templados. Sin embargo, el tipo de eutrofización que nos ocupa
es la denominada cultural, mucho más rápida y grave que la natural.
Las fuentes antrópicas de nutrientes pueden ser puntuales o difusas,
y de origen doméstico, industrial, agrícola o ganadero.
La eutrofización puede tener un origen natural, proceso lento, aunque
algo menos si se dan unas determinadas características geomorfológicas
en la cuenca (cuencos poco profundos y rocas no calizas), suelos ricos en nutrientes
y climas templados. Sin embargo, el tipo de eutrofización que nos ocupa
es la denominada cultural, mucho más rápida y grave que la natural.
Las fuentes antrópicas de nutrientes pueden ser puntuales o difusas,
y de origen doméstico, industrial, agrícola o ganadero.
Situación
actual y tendencias de la eutrofización de embalses en España
En el Libro Blanco del Agua en España (Ministerio de Medio Ambiente,
1998) se resume el estado trófico de los embalses españoles a
partir de información obtenida por los reconocimientos limnológicos
y estudios de teledetección del CEDEX entre 1990 y 1997.
Se ofrece en dicho
documento una panorámica bastante preocupante. Casi el 50% del volumen
de agua almacenado en los embalses españoles padece un estado "degradado"
(eutrófico o hipereutrófico). Los ámbitos de planificación
que presentan una mayor reserva degradada son las cuencas del Tajo (68%), Internas
de Cataluña (67%), Galicia Costa (64%) y Duero (57%). En la distribución
geográfica de los embalses se aprecia que es en los tramos bajos y medios
de los ríos, una vez superadas las grandes concentraciones urbanas, donde
la incidencia es mayor. Sin embargo, también habría que considerar
en la comparación la capacidad de los sistemas receptores y, en este
sentido, el Noroeste español y el Sistema Central tienen una propensión
mayor a la eutrofización, puesto que el sustrato silíceo de las
cuencas reduce la precipitación de fósforo con calcio y aumenta
su tiempo medio de residencia en la columna de agua. También hay que
tener en cuenta en la interpretación causal de esta distribución
que algunas de las principales ciudades costeras de España vierten directamente
al mar (en el mar Mediterráneo, históricamente oligotrófico,
la eutrofización ya es también un grave y acuciante problema).
Existen otros antecedentes también válidos en los estudios limnológicos
de los embalses españoles, recogidos en Álvarez Cobelas (1991),
que concluyen que el 50% de los embalses españoles son eutróficos
y el 20% hipereutróficos.
En aquellos en los que se dispone de contrastes recientes se evidencia un empeoramiento
progresivo, excepto en los que han alcanzado un umbral asintótico en
su grado trófico, determinado por los condicionantes físicos e
hidráulicos del sistema. Así por ejemplo, en ocho embalses de
la Cuenca Hidrográfica del Duero se ha realizado recientemente un completo
estudio de eutrofización (C.H.D., 1997) en el que se ha podido constatar
esta tendencia general. En los siete embalses en los que había catalogaciones
previas desde 1975, dos han evolucionado de mesotrofia a eutrofia, uno de oligotrofia
a mesotrofia, uno de oligotrofia a mesoeutrofia y los demás se han mantenido
en lo que parece su umbral máximo (mesotrofia, eutrofia o hipereutrofia,
según los casos).
El funcionamiento de los embalses como ecosistemas acuáticos
La
estructura física del medio
Los embalses constituyen elementos de retención hídrica en el
flujo unidireccional del río y añaden un desarrollo vertical al
sistema. A medida que se avanza desde la cola hacia la cabecera (presa), que
suele ser el punto de mayor profundidad, se pierden características fluviales
y se incrementa la similitud con los lagos. Sin embargo, se mantienen dos diferencias
transcendentales con los ambientes lacustres: los tiempos de retención
hidráulica son inferiores y las salidas de agua se producen en profundidad,
con un desacoplamiento más evidente del régimen hidrológico
en la cuenca de drenaje.
Si la profundidad es suficiente, en nuestro ámbito se desarrolla estacionalmente
una compartimentación física (estratificación térmica)
generada por un gradiente de densidad que implica una diferenciación
metabólica del sistema. Quiere esto decir que existen dos zonas claramente
diferenciadas y con un comportamiento ecológico divergente:
a. Una zona superior productora (epilimnion) en la que el tiempo de residencia
hidráulica, la provisión de nutrimentos y la entrada de energía
lumínica son suficientes para generar comunidades fitoplanctónicas
de entidad. En esta zona hay una producción neta de materia orgánica
y de oxígeno y mantiene intercambios con la atmósfera y con los
estratos inferiores.
b. Una zona inferior (hipolimnion) en la que predomina la respiración
sobre la producción y en la que se mineraliza materia orgánica
con un consumo de oxígeno. Mantiene un contacto activo con los sedimentos
y los flujos con el estrato superior (a través de una frontera de máximo
gradiente conocida como metalimnion) son predominantemente unidireccionales,
en el sentido de la fuerza de la gravedad.
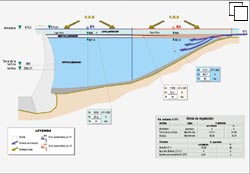 Fig.
1. El embalse de Buergomillodo, en le río
duratón, sirve para ilustrar la configuración característica
de un embalse que se estratifica en la época estival.
Fig.
1. El embalse de Buergomillodo, en le río
duratón, sirve para ilustrar la configuración característica
de un embalse que se estratifica en la época estival.
Hacer
clic sobre la imagen para verla ampliada
Esta descripción
básica (Fig. 1) presenta en la realidad numerosas modificaciones y situaciones
intermedias, pero en los embalses son muy relevantes los flujos hídricos
de entrada y salida cuyas características determinan los tiempos de retención
en los diferentes compartimentos, la carga de nutrientes que cada uno recibe,
su capacidad de asimilación y la carga evacuada. La configuración
concreta de esta compartimentación depende también en cada caso
de las características morfométricas del cuenco. En el ejemplo
de la figura 1 se aprecia una reserva hipolimnética grande en comparación
con la capa productora y el ingreso de nutrientes por el tributario principal
se produce en la capa profunda.
Interacciones
básicas en el sistema
Las comunidades biológicas que se desarrollan en el embalse se superponen
a este condicionado abiótico e interaccionan con él, de forma
que la resultante de estas interacciones determina la calidad del agua en cada
momento del ciclo anual.
Estas biocenosis se organizan en redes alimentarias con una estructura jerárquica
en niveles de organización que transitan por una escala temporal definida
por sus tiempos de generación y que transcurre, grosso modo, desde horas
en las bacterias, pasando por días en el fitoplancton, hasta semanas/meses
en los invertebrados y años en los peces. Cada nivel en estas redes integra
las tensiones que reciben del entorno, directamente o a través de los
demás niveles con los que interactúa. La subsecuente reacción
se expresa mediante mecanismos reguladores si no se sobrepasan los umbrales
de integridad ecológica; en caso contrario se producen colapsos, como
mortandades masivas, pérdidas de especies o, lo que es más grave,
de funcionalidad del nivel, lo cual puede acarrear profundas alteraciones en
el medio.
Considerando las interacciones directas e indirectas entre niveles, se puede
establecer las regulaciones principales de unos niveles sobre otros y determinar
aquellas que pueden resultar limitantes. Así resulta posible utilizar
este conocimiento para la gestión de la calidad del agua, puesto que
cada combinación de factores limitantes favorecerá la proliferación
de unas estrategias en detrimento de otras. Los organismos favorecidos modificarán
a su vez la combinación vigente de dichos factores, en un proceso dinámico
que define la sucesión ecológica en el sistema.
Los
ciclos de nutrientes
Los factores potenciales de limitación del crecimiento en cada nivel
son diversos (luz, hidrodinámica, nutrientes, depredación) y generalmente
varían a lo largo del año. A menudo, durante la primavera tardía
y el verano, es la provisión de nutrientes para los productores primarios
(fitoplancton en el caso de los embalses) el factor que predomina y define el
tipo de comunidad que se asienta, así como los flujos de materia y energía
hacia los niveles superiores -proceso conocido como regulación de abajo
hacia arriba-.
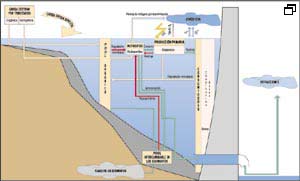 Fig.
2. En los ciclos de los nutrientes intervienen
decisivamente los compartimentos biológicos del embalse (véase
explicación en el texto).
Fig.
2. En los ciclos de los nutrientes intervienen
decisivamente los compartimentos biológicos del embalse (véase
explicación en el texto).
Hacer clic sobre la imagen para verla ampliada
En la figura 2 se representan aquellos compartimentos y flujos que pueden influir más directamente en el estado trófico de las aguas. La carga externa de nutrientes, que pueden ser orgánicos o inorgánicos, procede de los tributarios y de aportes directos por escorrentía y precipitación. Esta carga se reparte y procesa en el embalse principalmente en función de las variables hidrodinámicas y físicas (especialmente la temperatura) y alimenta a la reserva de nutrientes inorgánicos directamente o a través de procesos previos de mineralización.
Esta reserva de
nutrientes, en condiciones de intensidad y calidad lumínica (frecuencia
fotosintéticamente activa o PAR) suficientes, es utilizada por las algas
para su crecimiento.
El compartimento algal interacciona con el resto de las comunidades biológicas
del embalse, como se verá más adelante. Tanto el nitrógeno
como el carbono inorgánico y el oxígeno tienen una fase atmosférica
que, en el primer caso, está mediada por los procesos biológicos
de desnitrificación y fijación. Existen, por otra parte, múltiples
procesos de retorno a través del reciclaje directo de nutrientes o de
degradación y mineralización de los detritos. Estos pasos pueden
estar mediados también por los sedimentos, en los que existe una fracción
lábil de nutrientes que en determinadas condiciones retorna a la columna
de agua.
Cuando es la acción de un nivel superior sobre otro inferior (por ejemplo,
el pastoreo del zooplancton sobre el fitoplancton) lo que está limitando
el crecimiento poblacional, se habla de regulación de arriba hacia abajo.
En la medida en que se conozcan los elementos fundamentales de regulación
en cada caso, será posible intervenir mediante medidas que favorezcan
aquellos que limiten los procesos que llevan al colapso.
Uno de los elementos más frecuentemente limitante en los embalses es
el fósforo, de modo que la introducción de mayores cantidades
dispara el crecimiento algal hasta densidades muy elevadas en las que se pueden
imponer limitaciones de otros tipos (físicas y/o biológicas).
En una situación
muy característica en los embalses eutrofizados, se inicia en la primavera
una sucesión de comunidades de algas dominadas típicamente por
diatomeas pennadas que crecen en un medio abundante en recursos nutricios, de
modo que se pueden dar en este periodo los máximos anuales de biomasa
algal (generalmente estimada mediante la concentración de clorofila a).
Estos pulsos son seguidos de fases en las que se eleva la transparencia del
agua y abunda el zooplancton herbívoro y que anteceden al crecimiento
de las clorofíceas típico del verano temprano, cuando la energía
lumínica que entra en el sistema es máxima.
A medida que avanza el verano se puede producir una depleción de nutrientes
por el propio consumo algal que favorezca a otros grupos algales, como las diatomeas
céntricas, algunos géneros de crisofitas, de desmidiáceas
o de clorofíceas coloniales. Dependiendo del grado trófico y de
los nutrientes que limiten el crecimiento algal en este periodo, se verán
favorecidas diferentes estrategias al final del estiaje, desde cianofíceas
fijadoras de nitrógeno a otros tipos de cianofíceas, pasando por
dinoflageladas. En realidad, es la combinación de la limitación
por la energía y por los recursos la que determina la asociación
algal que se verá favorecida en cada momento (Reynolds, 1996).
En los embalses españoles, que abarcan un ámbito biogeográfico
de gran amplitud y diversidad, este cuadro simplificado se complica y las asociaciones
dominantes en cada caso atienden a tres factores de distribución básicos
(Riera et al., 1992):
a. El contenido mineral y la alcalinidad del agua.
b. El estado trófico del embalse.
c. La estabilidad de la columna de agua y el tiempo de residencia del agua.
Las redes alimentarias en los embalses poseen dos componentes claramente diferenciadas
(pelágica y bentónica) aunque interconectadas. La interconexión
principal se establece mediante la vía detrítica que enriquece
los fondos en materia orgánica y que se intensifica con la eutrofización.
A esta vía detrítica contribuye en gran medida el plancton, que
se constituye en un componente crucial y que además interesa controlar,
considerando los efectos perjudiciales de las proliferaciones masivas fitoplanctónicas.
A efectos tróficos, el fitoplancton es el principal, y en muchos embalses
casi el único, productor primario del sistema. Sin embargo, la eficiencia
y rutas con que esa producción se canaliza a otros componentes de la
red alimentaria es muy variable y depende del tamaño y digestibilidad
del tipo algal que se vea favorecido por los condicionantes ambientales concurrentes.
 |
En el conjunto del sistema, existen dos vías de consumo principales:
ingestión por los consumidores primarios (zooplancton) o evacuación
por el emisario. La producción no consumida ni evacuada se incorpora
directamente a la vía detrítica que, en gran parte, sedimenta.
Durante el proceso de mineralización de este componente detrítico,
protagonizado por las comunidades bacterianas del agua 2
y de los sedimentos, se consume oxígeno y se regeneran nutrientes inorgánicos
que pasan a disposición de los productores primarios, si se da la necesaria
coincidencia espacio-temporal.
Esta coincidencia está determinada por factores hidrodinámicos
y de equilibrios químicos de óxido-reducción; así,
por ejemplo, si durante el periodo de estratificación se produce un estado
de anoxia hipolimnética por una provisión excesiva de materia
orgánica en sedimentación desde la zona fótica superior,
los equilibrios en los sedimentos se desplazan hacia formas reducidas que previamente
han precipitado (principalmente el hierro) con los fosfatos; en estado reducido
son solubles y facilitan la reincorporación de los fosfatos a la columna
de agua en un proceso conocido como recarga interna de fósforo. Este
proceso frustra muy a menudo las expectativas de recuperación de la calidad
del agua en los embalses después de haber corregido las cargas externas
de fósforo.
Las clasificaciones taxonómicas constituyen un primer y necesario paso
en el procesamiento de la información sobre las biocenosis de los embalses,
cuando se requiere diagnosticar su estado y marcar unas pautas de gestión
de la calidad de las aguas. Sin embargo, es necesario llegar más allá
y, para esclarecer el funcionamiento del sistema, hay que asignar las categorías
taxonómicas (especies, géneros o familias, según los casos)
a grupos funcionales que constituyen nodos básicos en la red alimentaria
y/o grupos que representan un proceso crucial para la gestión (por ejemplo,
algas tóxicas).
 |
Así, una de las muchas clasificaciones del fitoplancton de embalses que pueden resultar útiles es la que se facilita en el cuadro 2. Los grupos funcionales responden también a criterios ecológicos: óptimos de crecimiento y umbrales de supervivencia según la disponibilidad de recursos nutricios y energéticos.
Zooplancton
Los componentes del zooplancton constituyen una compleja red que típicamente
se sostiene directa (pastoreo) o indirectamente (detritofagia) sobre la producción
fitoplanctónica en los embalses. Una de las clasificaciones funcionales
posibles, elaborada a partir de Armengol (1980), podría ser la que se
proporciona en el cuadro 3. No obstante, la dinámica poblacional de algunos
de estos componentes del plancton es muy compleja, con numerosos estadios del
desarrollo que mantienen estrategias ecológicas dispares entre sí.
En relación con los grupos de crustáceos planctónicos (copépodos y cladóceros), sí se ha comprobado su baja diversidad de especies respecto a otros países europeos con lagos. En este caso, los embalses introducen este tipo de hábitats pelágicos en España, país con escasez de lagos profundos naturales, y constituyen la base de una nueva colonización y diversificación biológica.
Bentos
En los embalses, los fondos suelen ser poco diversos y las comunidades litorales
tienen escasa representación debido a las fluctuaciones no sistemáticas
del nivel del agua. Esto implica una sensible reducción de la diversidad
de hábitats respecto a otros ecosistemas acuáticos, dado que el
medio bentónico (asociado a los fondos) es el que permite una mayor generación
de hábitats distintos. Por ello, las comunidades bentónicas de
los embalses suelen estar dominadas por escasos grupos, entre los que destacan
los gusanos anélidos oligoquetos y los quironómidos (larvas de
insectos). Sin embargo, estas asociaciones explotan el exceso de producción
primaria no utilizada y que se incorpora a la fracción detrítica,
de modo que pueden alcanzar considerables densidades.
Ictiofauna
Las especies de peces que colonizan los embalses españoles no están
adaptadas para explotar eficientemente las comunidades planctónicas,
de forma que sus dietas preferentes se encuentran en los detritos y faunas bentónicas.
No obstante, pueden consumir organismos del plancton de forma pasiva en función
de su densidad, de modo que cuando coinciden densidades planctónicas
altas en la zona fótica, por la que obligadamente transitan los peces
cuando hay anoxia hipolimnética, es más que probable que exista
un consumo apreciable de organismos del plancton. También hay que mencionar
el fitobentos de orilla, generalmente compuesto por microalgas en los embalses,
como una fuente de nutrimento para los peces. En todo caso, las dietas de la
ictiofauna en los embalses españoles son una cuestión que requiere
más investigación.
Las comunidades piscícolas en los embalses españoles están
dominadas por especies de elevada adaptabilidad a medios cambiantes (eurioicas),
en su mayor parte de la familia Cyprinidae. Esta familia, junto a los
Cobitidae, son las únicas exclusivas de agua dulce. La ictiofauna
epicontinental se completa con otras 10 familias, algunas de las cuales son
exóticas. En total, se han catalogado 61 especies, de las cuales 44 son
nativas y 17 exóticas (Granado Lorencio, 1996).
Aunque hay un considerable solapamiento de dietas entre especies y una variación
en el desarrollo de los hábitos alimentarios, se puede decir que, de
las especies más frecuentes en los embalses, la boga de río (Chondrostoma
polylepis) tiene hábitos más detritívoros, las carpas
(Cyprinus carpio) son muy adaptables y pueden explotar casi cualquier
recurso, y los barbos (Barbus sp.) son más especializados dentro
de un hábito omnívoro y explotan los invertebrados bentónicos
y el fitobentos. La resistencia a las bajas tensiones de oxígeno permite
a la carpa alcanzar recursos de los fondos hipolimnéticos durante el
estiaje, lo que les confiere una ventaja adaptativa evidente en los embalses
profundos y más eutrofizados.
En todo caso, parece que el grado trófico del agua está en relación
directa con la productividad de algunas de estas especies (Granado Lorencio
et al., 1985), a través de la productividad fitoplanctónica.
Entre la ictiofauna nativa existe un género (Alosa) que tiene
especialización planctívora aunque es una especie anfidroma (migra
entre el mar y el río); sin embargo, se conoce la existencia de poblaciones
"dulcificadas", es decir, que han quedado atrapadas en embalses y
completan su ciclo vital en él.
Una de las características de las comunidades ícticas de los embalses
españoles es la ausencia de depredadores con hábitos ictiófagos.
En la ictiofauna fluvial nativa es el grupo de los salmónidos el que
podría aportar esta función al sistema, pero sus requerimientos
ambientales son muy exigentes y estas condiciones sólo se presentan en
embalses de cabecera de cuenca. Sin embargo, los depredadores más frecuentes
en nuestros embalses pertenecen a especies exóticas, como el "black-bass"
(Micropterus salmoides), la lucioperca (Stizostedium lucioperca)
o el lucio (Esox lucius), aunque en este último caso hay controversia
acerca de su carácter exótico o nativo.
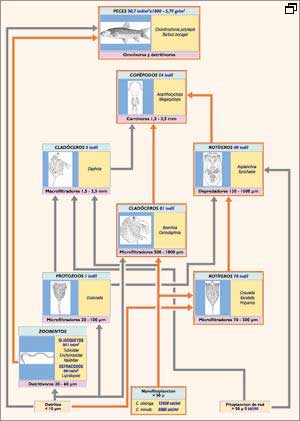 Fig.
3. Red alimentaria en un embálse hipereutrófico
del río Duero, en el inicio del período estival (véase
explicación en el texto). En rojo, elementos dominantes y flujos preferenciales
Fig.
3. Red alimentaria en un embálse hipereutrófico
del río Duero, en el inicio del período estival (véase
explicación en el texto). En rojo, elementos dominantes y flujos preferenciales
Hacer clic sobre la imagen para verla ampliada
En la figura 3 se representa, a modo de ejemplo, una red alimentaria establecida
para un embalse hipereutrófico, en la que se destacan en rojo las relaciones
tróficas más intensas. En este caso, el desmesurado crecimiento
algal debido a una provisión de nutrientes excesiva, procedente de aguas
residuales urbanas sometidas a tratamiento biológico, se traduce en densidades
muy altas y una dominancia casi exclusiva de algas de pequeño tamaño
del género Chilomonas. En estas condiciones, lo que resulta limitante
para el crecimiento algal es la provisión de luz por autoensombrecimiento
del fitoplancton, de modo que estas algas, que tienen flagelos para mantenerse
en la zona superior e incluso tienen un metabolismo mixto, por lo que pueden
extraer energía también a partir de compuestos orgánicos
(heterótrofos facultativos), encuentran una clara ventaja adaptativa.
Así se explica la abundancia relativa de microfiltradores en el zooplancton
que, a su vez, alimentan a una considerable población de copépodos
carnívoros que regulan su número y, por lo tanto, el consumo potencial
de algas. Las especies de peces presentes en el embalse explotan los recursos
bentónicos alimentados por la vía detrítica pero no contribuyen
a la reducción de la biomasa algal. A pesar de todo, se ha estimado que
una cantidad equivalente al 40% de la carga de fósforo anual que recibe
el embalse se encuentra en los peces, de forma que se convierten en una forma
potencial de extraer fósforo del sistema.
Las
posibilidades de la gestión limnológica
Resulta evidente en principio que la mejor técnica para evitar la eutrofización
de los embalses es corregir las fuentes de nutrientes, especialmente de fósforo,
en origen. Sin embargo, esto no es siempre posible desde el punto de vista logístico
o financiero.
En cualquier caso,
existen soluciones de tratamientos blandos que no están suficientemente
contrastadas y explotadas en nuestro país. En primer lugar, se ha de
atender a la cuenca vertiente en su conjunto y analizar todas las posibles medidas
que se pueden adoptar en los puntos críticos. Así, las alternativas
no se agotan en la adaptación de estaciones depuradoras convencionales
con mecanismos para la reducción de nutrientes, sino que se han de valorar
también las posibilidades de corrección en las actividades productivas.
A este respecto, la polémica surgida en Estados Unidos con la sustitución
de los fosfatos en los detergentes es aleccionadora (Edmondson, 1991). La importancia
crucial de controlar el ingreso de fosfatos en el medio no es ajena tampoco
a las administraciones europeas, como lo refleja la existencia de un centro
de investigación dedicado a esta cuestión. 3
Aun así, todavía caben muchas opciones entre los vertidos
y el embalse, que tienen la ventaja de que permiten tratar las aportaciones
de carácter difuso. Se citan, por ejemplo, la rehabilitación y/o
construcción de humedales, la reforestación de las cuencas, en
especial, de la vegetación de ribera, la construcción de plantas
de reducción de fósforo en los tributarios, los preembalses, la
recarga de acuíferos y la optimización del hábitat fluvial
para la retención de nutrientes.
Por otro lado y como se ha visto, las características del embalse y su
régimen de explotación modulan su respuesta a una carga de nutrientes
dada. Esto significa que existe un cierto margen de intervención mediante
la modificación de determinados procesos en el propio embalse.
Así, se pueden citar como posibilidades genéricas las siguientes
(modificado a partir de Straskraba, 1993):
a. Regulación hidráulica: permite modificar las tasas de renovación
de los diferentes compartimentos, según interese en cada caso y momento
del año. Sus posibilidades dependen de la configuración de las
estructuras de salida de la presa y de las demandas aguas abajo.
b. Aireación y mezcla de la columna de agua: se refiere a métodos
que buscan modificar la hidrodinámica y la compartimentación física
del sistema (desestratificación mecánica o neumática mediante
aireación) o combatir el déficit hipolimnético de oxígeno
sin deshacer dicha compartimentación (aireación u oxigenación).
c. Inactivación del fósforo: medidas de tipo físico y químico
dirigidas a reducir la biodisponibilidad del fósforo en el sistema. La
coagulación y precipitación química con alúmina
es la más utilizada.
d. Tratamiento de los sedimentos: incluye posibilidades como la cubrición
de los sedimentos con materiales inertes o la remoción y aireación
de los sedimentos.
e. Modificación del ambiente lumínico: hace referencia a la reducción
de la intensidad de luz mediante la provisión de sombra, la suspensión
de partículas o la tinción directa del agua.
f. Biomanipulación: esta familia de técnicas se refiere tanto
a la eliminación directa de las algas mediante tóxicos selectivos
que, en general, no es muy recomendable, como a la gestión de las comunidades
piscícolas del embalse para controlar el zooplancton y, a su vez, reducir
la biomasa algal.
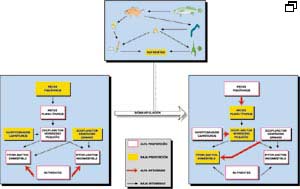 Fig.
4. Esquema que representa las modificaciones
en la red trófica que se persiguen con las estrategias de biomanipulación.
Fig.
4. Esquema que representa las modificaciones
en la red trófica que se persiguen con las estrategias de biomanipulación.
Hacer
clic sobre la imagen para verla ampliada
La biomanipulación para controlar la biomasa algal desde los eslabones
tróficos superiores (Fig. 4) consiste en incrementar la presión
sobre los peces planctívoros, bien mediante la introducción de
depredadores o bien mediante la extracción directa. De esta forma, aumentará
la biomasa de zooplancton herbívoro grande, que puede mantener así
la densidad de las algas comestibles en determinados niveles.
Se pueden incluir
también en este apartado las estrategias de gestión piscícola
encaminadas a optimizar la incorporación de fósforo a la biomasa
de peces y la extracción de la misma.
Como se puede apreciar, las posibilidades de gestión son diversas y las
medidas que se adopten deben integrarse en programas de recuperación
definidos a la escala de cuenca. Para ello, se requiere la obtención
de dos tipos de información complementaria:
a. Información a largo plazo de fuentes y procesos clave en la cuenca
y embalses, que permita seguir la evolución interanual de la problemática
y desarrollar y ajustar modelos empíricos sencillos para generar esquemas
de decisión en la planificación a escala regional.
b. Información intensiva en los embalses, en uno o dos ciclos anuales,
para establecer los balances internos y el funcionamiento limnológico
del sistema con una resolución espacio-temporal adecuada a los patrones
de distribución y a las tasas de renovación de cada nivel de organización
tratado. Con esta información se pueden establecer modelos biogeoquímicos
del sistema con suficiente detalle como para habilitar la toma de decisiones
a nivel local. Estas actuaciones estarían encaminadas a recuperar una
calidad de las aguas suficiente para mantener el sistema dentro del rango de
integridad ecológica.
Conclusión
A medida que se va dando solución a los problemas más acuciantes
de la calidad del agua, se van evidenciando nuevos problemas que tienen un carácter
menos agudo para las poblaciones humanas, es decir, que no suelen causar mortalidad
directa e inmediata, pero con un alcance espacio-temporal mayor, lo que significa
que su reversión es más lenta y compleja. Así, por ejemplo,
en el siglo XIX los problemas de calidad se presentaban como septicemias epidémicas
locales de carácter anual; ya en el siglo XX, ha preocupado inicialmente
la contaminación orgánica por aguas residuales; en los años
setenta la contaminación térmica, la eutrofización y la
contaminación por sustancias tóxicas; en los años ochenta,
la contaminación por nitratos, acidificación, contaminación
accidental; y en la última década el calentamiento global y cambio
global, que son fenómenos de alcance planetario y de escala temporal
milenaria.
La eutrofización cultural constituye un proceso contaminante de alcance
regional y que se desarrolla en una escala de tiempo que se cifra en lustros
o en décadas. Por ello, se están empezando a validar ahora modelos
propuestos en los años ochenta (OECD, 1982), pero que apenas tratan los
compartimentos biológicos. La respuesta de estos compartimentos carece
aún de suficiente base empírica y los embalses proporcionan una
ocasión única para definirla cuantitativamente.
Por otra parte, resulta evidente el interés que tiene mantener el sistema
en unas condiciones de calidad que se muevan dentro de los márgenes de
la integridad ecológica, es decir, en los que se minimizan los riesgos
de aparición de situaciones indeseables. Los episodios de escasez de
agua, que en nuestro país son frecuentes, han permitido constatar que
la provisión del recurso a la sociedad no es sólo un problema
de cantidad sino también de calidad.
El beneficio de esa gestión sostenible no estriba sólo en lo socioeconómico,
en cuanto a garantizar las demandas de agua, sino que es también ecológico,
puesto que se mejoran las condiciones de conservación de las especies
acuáticas y se aumenta la biodiversidad; además, se frena la propagación
del problema a ecosistemas fluviales y también estuarinos situados aguas
abajo.
Por otro lado, no se está atendiendo suficientemente a las nuevas tecnologías
que están surgiendo para la corrección de estos problemas de contaminación
de las aguas. Este tipo de técnicas de la ecotecnología, que utilizan
en su beneficio conceptos de funcionamiento y procesos de los propios ecosistemas,
suelen representar alternativas de bajo coste y de menores efectos ambientales
colaterales. Sin embargo, su diseño no se está optimizando al
ritmo que podría hacerlo porque no hay un flujo de información
específica de nuestra geografía suficiente como para definir y
calibrar los modelos matemáticos del funcionamiento de las variables
de diseño. Digamos que es muy difícil mejorar lo que no se utiliza.
No obstante, existen algunas iniciativas de soluciones alternativas que se han
puesto en práctica en embalses españoles. En la monografía
«Embalses y medio ambiente» (Ministerio de Medio Ambiente, 1996),
se recoge la mayor parte de estas experiencias.
En cualquier caso, la problemática concreta de la eutrofización
de embalses no es banal, como se ha intentado reflejar aquí, y las escalas
espacio-temporales que implica recomiendan la instauración de mecanismos
que permitan una gestión continua y estable en el tiempo. Asimismo, constituye
un ejemplo claro de la multiplicidad de disciplinas que requiere el desarrollo
de modelos operativos del sistema, que debe implicar a técnicos de diversa
filiación, y de la importancia que tiene el disponer de información
suficiente de todos los compartimentos y procesos afectados.n
Notas
1. "Propuesta modificada de Directiva del Consejo por la
que se establece un Marco Comunitario de Actuación en el Ámbito
de la Política de Aguas. Posición Común. Documento del
2 de marzo de 1999".
2. A efectos de simplificación, las bacterias se suelen
incluir también en el propio compartimento de los detritos cuando la
información de campo es insuficiente para asignarles explícitamente
grupos ad hoc.
3. CEEP (Centre Européen d'Etude des Polyphosphates).